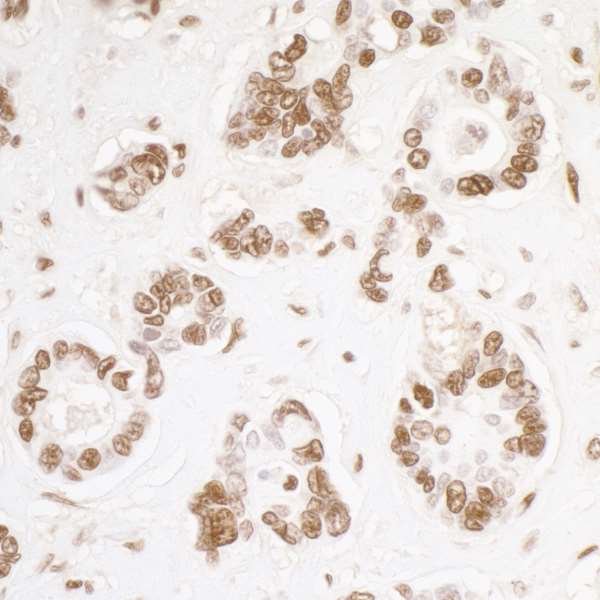
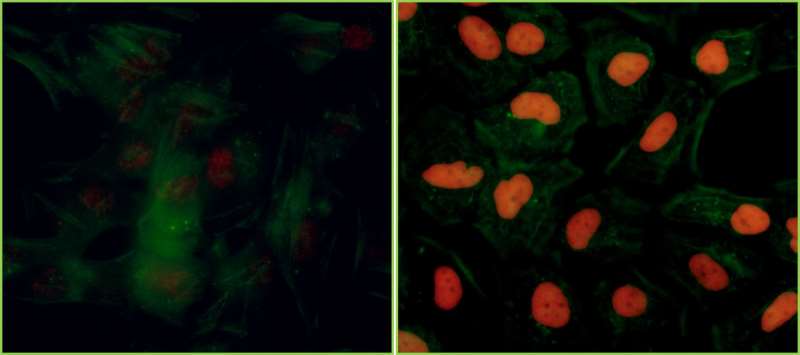
Central to KAP-1’s role in these processes is its ability to influence epigenetic patterns and chromatin compaction, thereby regulating the dynamic organization of chromatin structure.1 These and other roles of KAP-1 are mediated through its function as an interacting protein for KRAB zinc finger proteins (KRAB-ZFPs). The conserved KRAB repression domain is present in many transcription factors. Binding of KAP-1 to this domain results in transcriptional co-repression.3 The action of KAP-1 is not restricted to its interaction with KRAB-ZFPs, however, in that transcription factors that lack the KRAB-ZFP domain can also be regulated by KAP-1; examples of such transcription factors include c-Myc and E2F transcription factor 1 (E2F1).4,5
KAP-1 and other co-repressors may act through 1) recruitment of enzymes that modify histones by addition or removal of acetyl groups; 2) regulation of nucleosome spacing by modulation of ATP-dependent nucleosome-remodeling enzymes such as chromodomain helicase DNA binding protein 3 (CHD3) and 4 (CHD4); or 3) stabilization of chromatin by recruitment of specific proteins such as heterochromatin protein (HP1), which binds to methylated histones and maintains their closed state. In any of these mechanisms, transcriptional silencing can be achieved by compaction and/or stabilization of chromatin to form heterochromatin.3,6
Regulation of the gene repressive function of KAP-1 is coordinately achieved through post-translational modification of KAP-1, namely via sumoylation or phosphorylation.1-3,6 Indeed, the repressive capacity of KAP-1 is controlled by its existence in either a sumoylated or phosphorylated state, in that sumoylated and phosphorylated KAP-1 are involved in transcriptional repression and DNA repair, respectively.7
Under normal conditions, KRAB-ZFPs recruit sumoylated KAP-1 to the genome. Upon DNA damage, robust and transient ATM serine/threonine kinase-mediated phosphorylation of KAP-1 occurs. Subsequently, KAP-1 is rapidly localized to the area of DNA damage, where it may facilitate localized chromatin decondensation. This decondensation in turn allows access of DNA repair proteins to the area of damage. Following DNA damage repair, the chromatin may be re-condensed following KAP-1 sumoylation.1
At present, it’s clear that the function of KAP-1 is highly dependent on post-translational modifications; sumoylated KAP-1 is involved in transcriptional repression, whereas phosphorylated KAP-1 is involved in DNA repair. Future work will no doubt continue to elucidate the diverse cellular processes in which KAP-1 plays a role and the mechanisms underlying these influences.